| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
| |
|
一切恩爱会,无常难得久;
生世多畏惧,命危于晨露。 由爱故生忧,由爱故生怖; 若离于爱者,无忧亦无怖。 |
|
 汇总4月最新:临床试验招募| 小细胞
"化疗±免疫后进展了,还能怎么办?"——这是许多小细胞肺癌患者和家属的困境。
临
汇总4月最新:临床试验招募| 小细胞
"化疗±免疫后进展了,还能怎么办?"——这是许多小细胞肺癌患者和家属的困境。
临
 ALK+肺癌患者眼神里的光,正在被看见
今天,上海的夏日阳光格外明亮。
在上海市徐汇区建国西路386号甲,一场属于ALK+非小细
ALK+肺癌患者眼神里的光,正在被看见
今天,上海的夏日阳光格外明亮。
在上海市徐汇区建国西路386号甲,一场属于ALK+非小细
 求助各位老师,肺腺癌晚期无靶点一线
2025-8-12 术前评估早期,术中发现胸膜播散,切除原发灶
手术医院病理:大细胞癌,未
求助各位老师,肺腺癌晚期无靶点一线
2025-8-12 术前评估早期,术中发现胸膜播散,切除原发灶
手术医院病理:大细胞癌,未
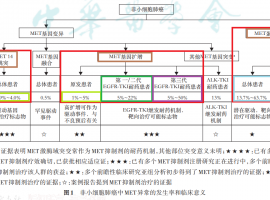 奥西伏美阿美耐药怎么办?基因全因没
你以为我想盲试?,这不也是没有办法的办法?谁不想正规治疗,按照指南和循证医学的节
奥西伏美阿美耐药怎么办?基因全因没
你以为我想盲试?,这不也是没有办法的办法?谁不想正规治疗,按照指南和循证医学的节
 塔拉妥单抗获批!小细胞肺癌治疗迎来
作者:seacat今年2月份举行的第23届中国肺癌高峰论坛主题是 “历史转折关头的小细胞肺
塔拉妥单抗获批!小细胞肺癌治疗迎来
作者:seacat今年2月份举行的第23届中国肺癌高峰论坛主题是 “历史转折关头的小细胞肺






 寄予极大期望的易要不要再坚持吃段时间?才7天,可是感觉每天都在加重病情。明明,免疫组化是中阳啊!为什么要去测该死的突变,导致犹豫着始终不敢用。。是扩散太多了吗?失控了吗?感觉左眼周似乎有小的丘疹出现?加量吃么?还是换299804+184?
寄予极大期望的易要不要再坚持吃段时间?才7天,可是感觉每天都在加重病情。明明,免疫组化是中阳啊!为什么要去测该死的突变,导致犹豫着始终不敢用。。是扩散太多了吗?失控了吗?感觉左眼周似乎有小的丘疹出现?加量吃么?还是换299804+184? 显身卡
显身卡




